¶ Inhibidores de la Bomba de Protones
Carrera de Medicina – Facultad de Ciencias Químicas y de la Salud (UTMACH)
Autores: Nayerli M. Miranda R.; María M. Hurel R.; Lesly M. Orellana L.
Tutor: Dr. Roberto Eduardo Aguirre Fernandez, PhD.
¶ Introducción
El estómago es un órgano encargado de almacenar, mezclar y descomponer los alimentos, además de secretar HCl y enzimas digestivas que facilitan la digestión y eliminación de microorganismos [1]. Su actividad está regulada por el SNC mediante mecanismos neuroendocrinos y autonómicos, especialmente a través del nervio vago, que libera acetilcolina y estimula la secreción gástrica [2].
La célula parietal es el principal responsable de la producción de HCl mediante la acción de la bomba de protones que se encuentra ubicada en su membrana apical [3]. La secreción ácida está modulada por receptores estimuladores e inhibidores; por tanto, si existen alteraciones en este equilibrio, pueden generarse trastornos ácidos–pépticos [4].
¶ Desarrollo
El ácido clorhídrico (HCl) desempeña un papel crucial en la digestión de los alimentos, permitiendo la descomposición de nutrientes y actuando como una barrera protectora frente a patógenos ingeridos [5]. Este proceso es mediado por un grupo especializado de células gástricas, principalmente las células parietales, que secretan ácido en respuesta a diversos estímulos [6]. La secreción de ácido está regulada por una compleja red de vías neuronales, hormonales y paracrinas, que actúan en un sistema altamente coordinado [7]. Trastornos como la enfermedad por reflujo gastroesofágico (ERGE) y las úlceras gástricas han impulsado el uso de fármacos como los inhibidores de la bomba de protones (IBP), los cuales modulan esta secreción para tratar enfermedades relacionadas con la hiperacidez [8].
¶ Células gástricas involucradas en la producción de ácido
La mucosa gástrica tiene varias células especializadas que desempeñan funciones clave en la producción de ácido y en la protección del estómago. Entre las principales células involucradas en la secreción de ácido se encuentran las células parietales, las células principales, las células enterocromafines-like (ECL) y las células G [9].
¶ Células parietales
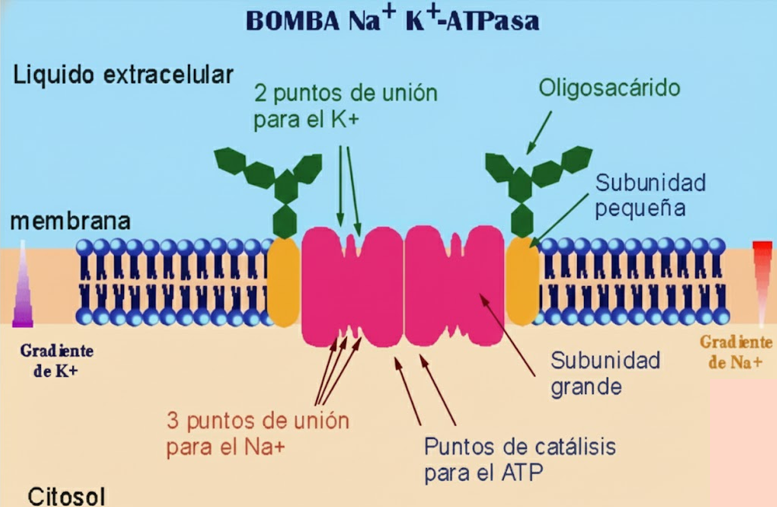
Las células parietales, también conocidas como células oxínticas, son responsables de la secreción de HCl. Estas células se localizan en el cuerpo y el fundus del estómago, y su principal función es acidificar el contenido gástrico para facilitar la digestión de los alimentos y destruir patógenos ingeridos [10]. La secreción de HCl se lleva a cabo a través de la H⁺/K⁺-ATPasa, que intercambia protones por iones potasio, a expensas de ATP; esta bomba se encuentra en la membrana apical de las células parietales y su actividad está regulada por diversos factores [11].
Vías de regulación:
- Histamina: La histamina es liberada por las células ECL y actúa sobre los receptores H₂ en las células parietales. La activación de estos receptores aumenta los niveles de cAMP en el interior de la célula, lo que a su vez activa la proteína quinasa A (PKA). Esta activación desencadena la translocación de los vesículos tubulovesiculares (TVs), que contienen H⁺/K⁺-ATPasa, hacia la membrana apical, incrementando la secreción de ácido [12].
- Gastrina: La gastrina, un péptido liberado por las células G, estimula la liberación de histamina desde las células ECL y también puede actuar directamente sobre las células parietales, potenciando la secreción de HCl [13].
- Acetilcolina: La acetilcolina, liberada por el nervio vago, actúa sobre los receptores muscarínicos M3 en las células parietales, lo que incrementa la liberación de calcio intracelular y estimula la secreción de ácido de manera independiente de la histamina [14].
¶ Células principales
Las células principales, localizadas en las glándulas gástricas del cuerpo y el fundus del estómago, se encargan de la secreción de pepsinógeno, un precursor inactivo de la pepsina, que es esencial para la digestión de proteínas [15]. Aunque no están directamente involucradas en la producción de ácido, su función se ve complementada por la acidez del estómago, que activa el pepsinógeno en pepsina [16].
¶ Células enterocromafines-like (ECL)
Las células ECL se encuentran en la mucosa gástrica y son responsables de la liberación de histamina. La histamina actúa como un potente secretor para las células parietales al activar los receptores H₂ y aumentar la secreción de ácido [17].
¶ Células G
Las células G, localizadas principalmente en el antro gástrico, producen gastrina en respuesta a la distensión del estómago o la presencia de proteínas; esta es la principal hormona que regula la secreción de ácido gástrico y actúa sobre los receptores CCK₂ de las células parietales y las células ECL; la gastrina estimula la liberación de histamina y la secreción de HCl [18].
¶ Regulación de la secreción gástrica
La secreción gástrica se controla por mecanismos nerviosos, hormonales y paracrinos, que actúan tanto para estimular como para inhibir la producción de ácido clorhídrico y otros componentes del jugo gástrico [19].
¶ Estimulación de la secreción gástrica
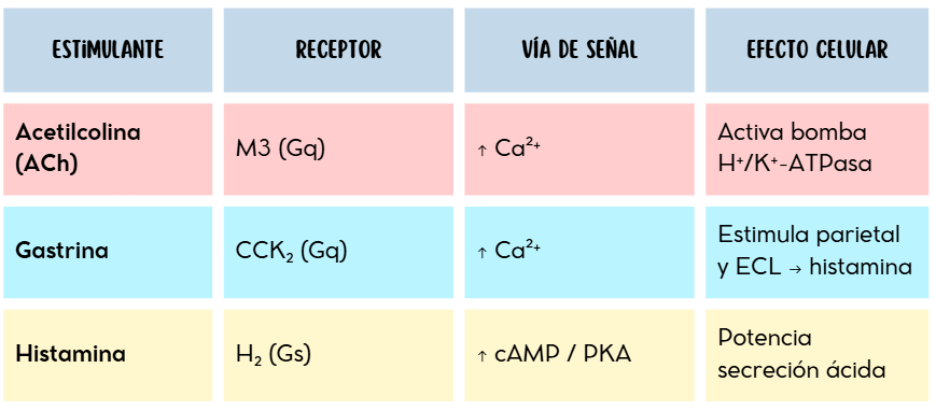
- La acetilcolina (ACh), liberada por terminaciones del nervio vago, se une a receptores muscarínicos M3 en las células parietales, activando la vía Gq/PLC/Ca²⁺, lo que estimula la bomba H⁺/K⁺-ATPasa y aumenta la secreción ácida [20].
- La histamina, liberada por las células enterocromafines (ECL), actúa sobre receptores H₂ (Gs), incrementando cAMP y potenciando la secreción inducida por gastrina y acetilcolina [21].
- La gastrina, secretada por las células G del antro gástrico, se une a receptores CCK₂ en las células parietales, también activando la vía Gq/Ca²⁺ y estimulando la liberación de histamina en las ECL [22].
¶ Inhibición de la secreción gástrica
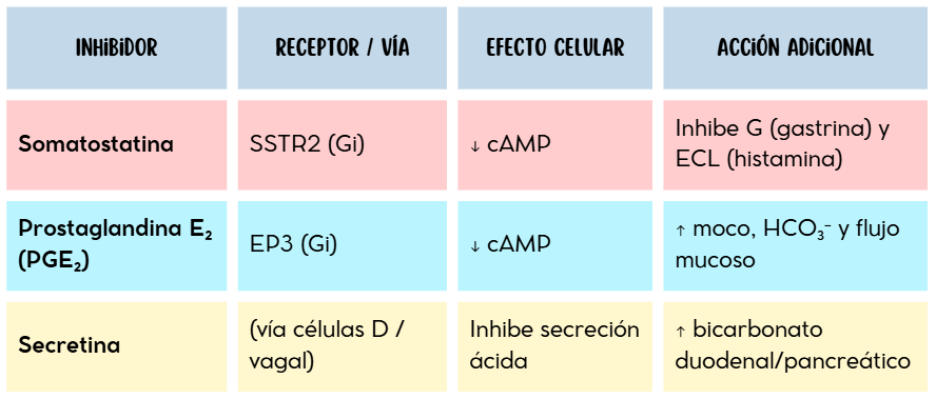
- Las prostaglandinas E₂ (PGE₂) ejercen un efecto protector e inhibidor sobre la secreción ácida al activar receptores EP3 (Gi) que reducen el cAMP y estimulan la producción de moco, bicarbonato y el flujo sanguíneo mucoso [23].
- La somatostatina, secretada por las células D, inhibe directamente la secreción de ácido mediante los receptores SSTR2 (Gi) y suprime la liberación de gastrina e histamina [24].
- La secretina, liberada por las células S del duodeno en respuesta a un pH ácido, estimula la secreción de bicarbonato pancreático y duodenal, y a su vez inhibe la secreción gástrica mediante la activación de somatostatina y reflejos vagales [25].
- El péptido inhibidor gástrico (GIP), aunque inicialmente se consideró un inhibidor de la secreción ácida, hoy se reconoce principalmente por su función incretínica; su papel en la inhibición gástrica es débil y variable en humanos [26].
¶ Mecanismo de retroalimentación (feedback)
La retroalimentación negativa regula la secreción de ácido según el pH intragástrico. Cuando el pH desciende por debajo de 3, se estimula la liberación de somatostatina desde las células D, que inhibe las células G (gastrina), ECL (histamina) y parietales, reduciendo así la secreción de ácido. Por el contrario, durante la ingesta de alimentos, los péptidos y aminoácidos amortiguan el pH, reducen la actividad de las células D, e inducen la secreción de gastrina, reactivando el circuito secretor [27].
¶ Receptores de la célula parietal
Las células parietales gástricas, como ya se mencionó anteriormente, se encuentran en las glándulas oxínticas del cuerpo y fundus del estómago, en las cuales son las encargadas de secretar ácido clorhídrico [28]. Su actividad está regulada por la interacción entre receptores estimuladores e inhibidores que responden a señales nerviosas, endocrinas y paracrinas [29]. Este equilibrio entre ambos tipos de receptores garantiza una adecuada digestión y la protección de la mucosa gástrica de la autolesión por exceso de ácido [30].
¶ Receptores estimuladores de la célula parietal y su acción sobre la bomba de protones
La estimulación de la célula parietal depende de 3 vías que son: nerviosa, hormonal y paracrina, las cuales actúan de forma sinérgica sobre receptores específicos [31]. Estos receptores activan mecanismos intracelulares distintos, pero todos convergen en activar la bomba de protones, que se encarga de secretar iones hidrógeno al lumen gástrico [32].
Los receptores estimuladores que encontramos en la célula parietal son: receptores muscarínicos M3, receptores histamínicos H₂ y receptores de gastrina (CCK₂) [33].
1. Receptores muscarínicos M3
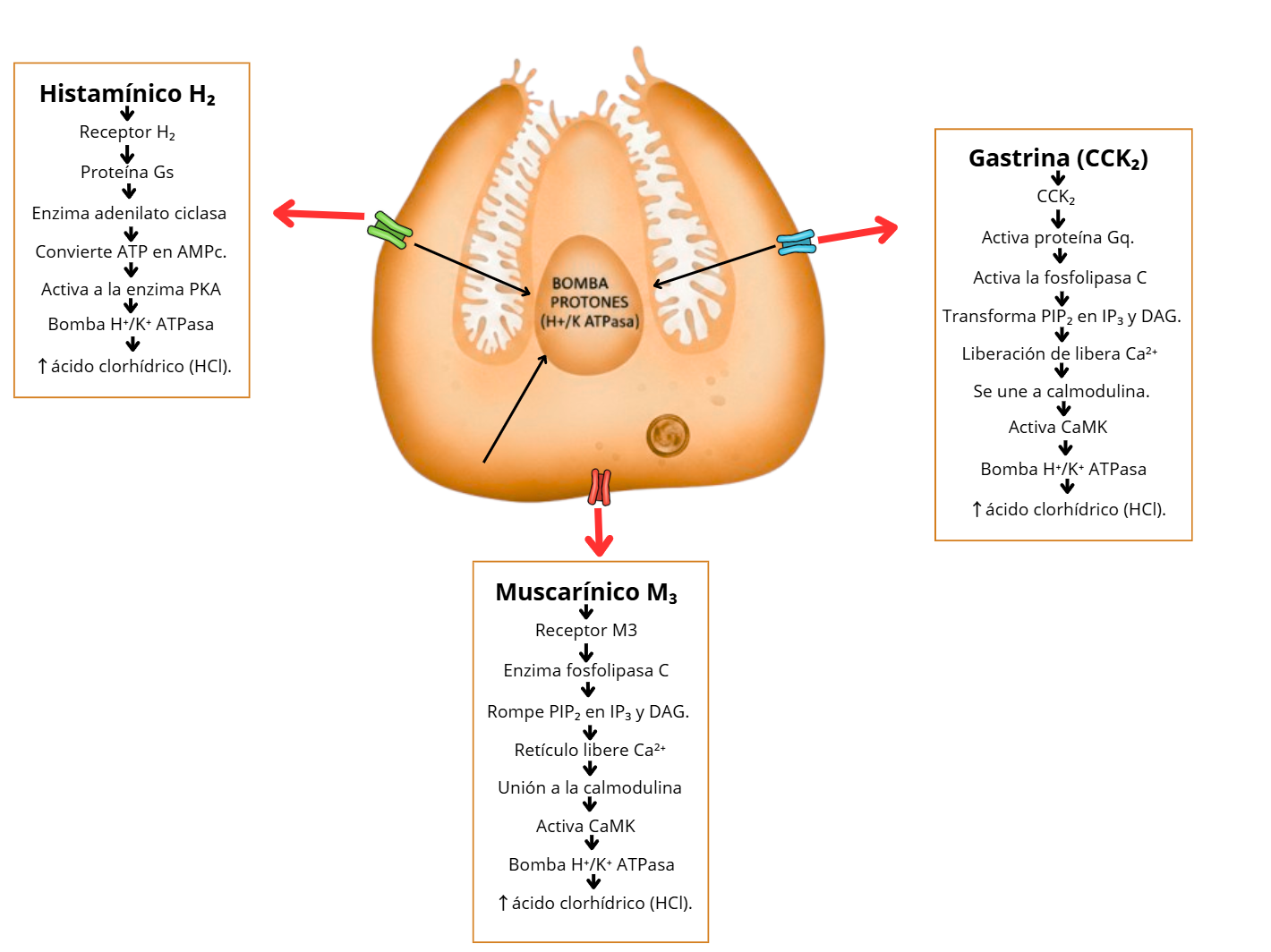
El sistema nervioso central regula la secreción gástrica por medio del nervio vago, que libera acetilcolina cuando inicia la digestión [34]. Esta acetilcolina se une a los receptores muscarínicos M3 que se encuentran ubicados en la membrana basolateral de la célula parietal [35]. Al unirse la ACh al receptor M3 se activa la proteína G del tipo Gq que estimula la fosfolipasa C; esta enzima transforma un fosfolípido de la membrana en dos segundos mensajeros: inositol trifosfato (IP₃) y diacilglicerol (DAG) [36].
El IP₃ provoca la liberación de calcio desde los depósitos intracelulares, y el DAG, junto con el aumento del Ca²⁺, activa a la proteína quinasa C (PKC) [37].
Ambas moléculas, es decir, Ca²⁺ y PKC, inducen la fusión de las vesículas intracelulares que contienen las bombas de protones con la membrana apical de la célula parietal, lo que provoca que la bomba se active y comience a expulsar protones hacia el lumen gástrico, intercambiándolos por iones de potasio [38].
2. Receptor histamínico H₂
La histamina es liberada por las células enterochromafines que se encuentran cerca de las células parietales; su función es actuar de manera paracrina, es decir, local, sobre las células vecinas [39].
Cuando la histamina se une al receptor H₂ en la célula parietal, este activa una proteína Gs que estimula la enzima adenilato ciclasa, la cual convierte ATP en AMP cíclico (AMPc) [40].
El aumento del AMPc activa la proteína quinasa A, la cual fosforila proteínas específicas dentro de la célula, desencadenando la movilización de las vesículas con bombas H⁺/K⁺-ATPasa hacia la superficie celular y, por tanto, aumentando la actividad de la bomba; el resultado es incrementar sostenidamente la secreción de ácido clorhídrico [41].
3. Receptor de gastrina (CCK₂)
La gastrina es una hormona secretada por las células G del antro gástrico en respuesta a la distensión del estómago o la presencia de alimento; esta gastrina viaja por la sangre hasta las células parietales, donde se une a receptores CCK₂ [42].
Su mecanismo de acción es similar al de la acetilcolina, ya que también utiliza una proteína G del tipo Gq, activando la fosfolipasa C que genera IP₃ y DAG, elevando el Ca²⁺ intracelular y activando PKC, lo que provoca la activación de la bomba de protones y, por tanto, el aumento de la secreción de HCl [43].
Además, la gastrina estimula de manera indirecta la secreción ácida al unirse también a receptores CCK₂ de las células enterochromafines, promoviendo la liberación de histamina que potencia aún más la acción sobre la célula parietal [44].
¶ Receptores inhibidores de la célula parietal y su acción sobre la bomba de protones
La secreción ácida del estómago no solo depende de señales estimuladoras, sino también de mecanismos inhibitorios que evitan la producción excesiva de HCl y protegen la mucosa gástrica [45]. Estos receptores inhibidores actúan directamente sobre la célula parietal o de forma indirecta sobre otras células que participan en la secreción ácida, como las células G y las células enterochromafines [46].
1. Receptores de somatostatina
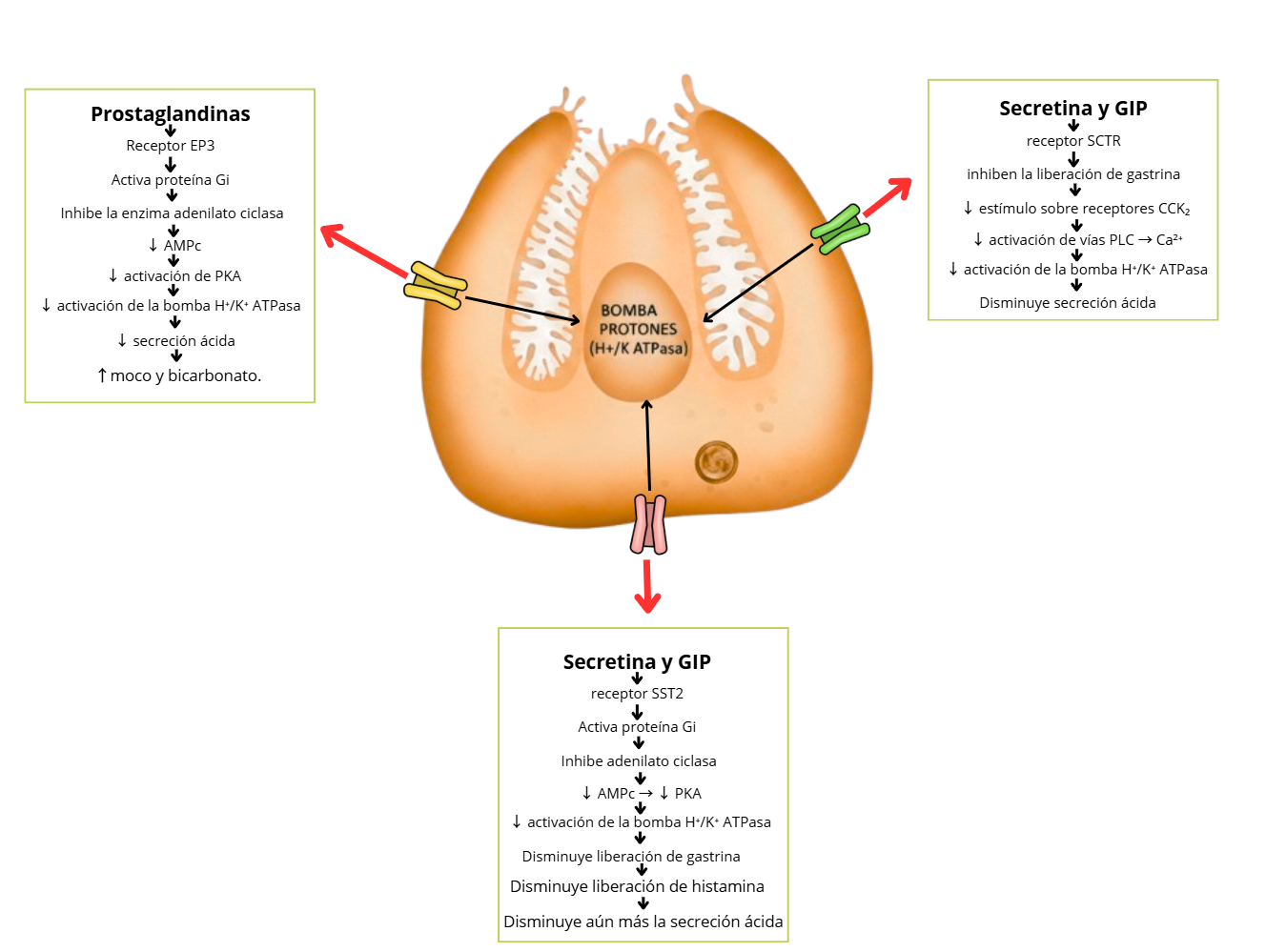
La somatostatina es una hormona producida por las células D, localizadas en el antro y cuerpo gástrico; actúa como un potente inhibidor paracrino y endocrino de la secreción ácida [47]. Cuando la concentración de ácido en el estómago aumenta, las células D liberan somatostatina, la cual se une a los receptores de somatostatina tipo 2 presentes en las células parietales [48].
Su efecto es inhibir el adenilato ciclasa, generando disminución de los niveles intracelulares de AMPc; dado a eso, sin AMPc, la proteína quinasa A no puede fosforilar las proteínas necesarias para movilizar la bomba de protones hacia la membrana, por lo que la secreción de ácido disminuye [49].
Cabe mencionar que la somatostatina tiene efectos indirectos sobre las células G al inhibir su acción y, por tanto, reducir la liberación de gastrina, y también sobre las células enterochromafines al reducir la liberación de histamina [50].
2. Receptores de prostaglandinas
Las prostaglandinas, sobre todo las PGE₂, son compuestos lipídicos sintetizados en la mucosa gástrica a partir del ácido araquidónico mediante la enzima ciclooxigenasa [51]. Estas sustancias se unen a los receptores de prostaglandinas tipo EP3 de la célula parietal, los cuales están acoplados a una proteína Gi; al activarse, inhiben el adenilato ciclasa, reduciendo los niveles de AMPc y, en consecuencia, la actividad de la bomba de protones [52].
Adicionalmente, el papel de las prostaglandinas no solo se limita a lo anterior, sino que también estimulan las células epiteliales del moco gástrico para producir moco y bicarbonato, creando una barrera protectora frente al ácido [53].
3. Receptor del péptido inhibidor gástrico (GIP)
El GIP es una hormona dependiente de la glucosa, producida por las células K del intestino delgado; su función es inhibir la secreción ácida y reducir la motilidad gástrica cuando el quimo pasa al intestino [54].
El GIP se une a receptores específicos en las células G y parietales, generando un bloqueo indirecto sobre la secreción de gastrina y reduciendo la actividad de la bomba de protones [55].
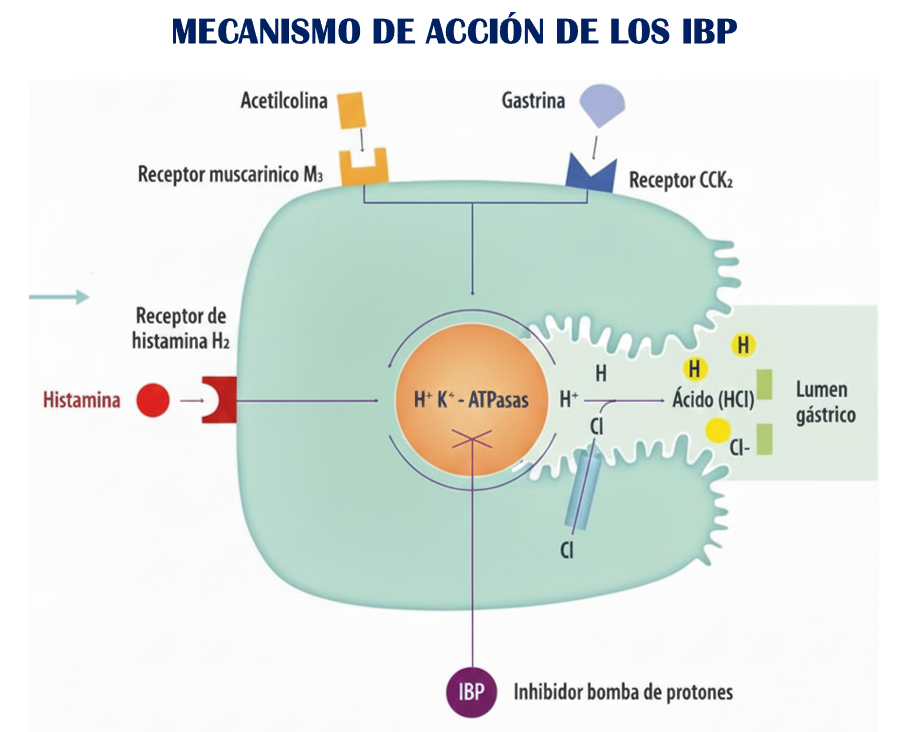
¶ Mecanismo de acción de los fármacos antisecretores IBP
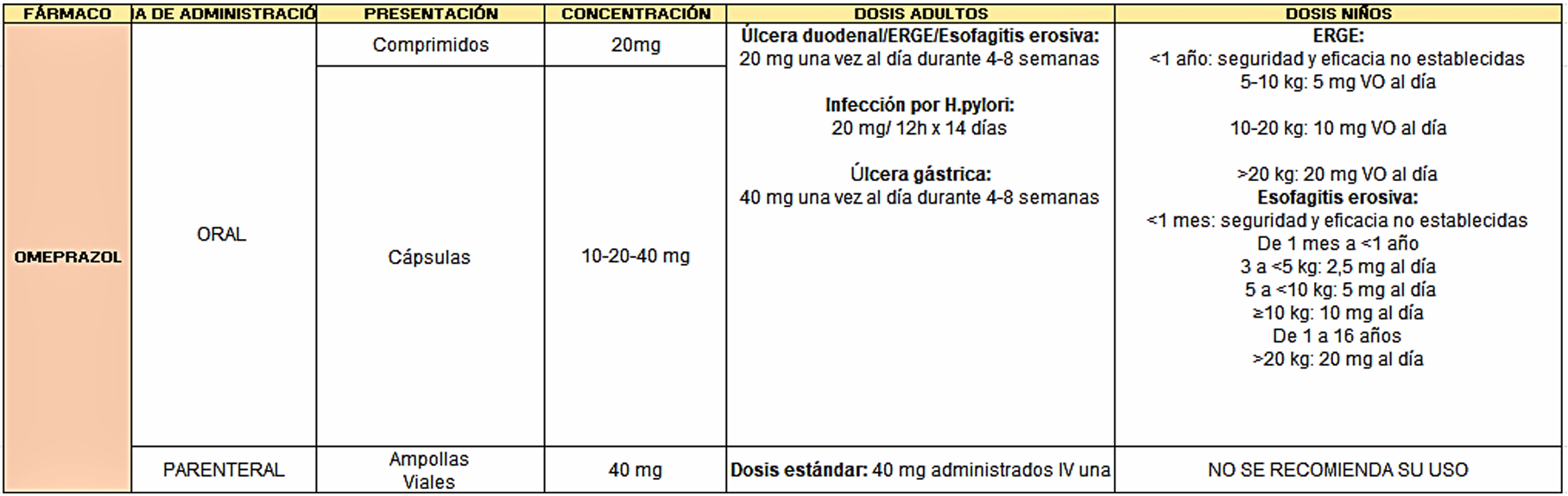
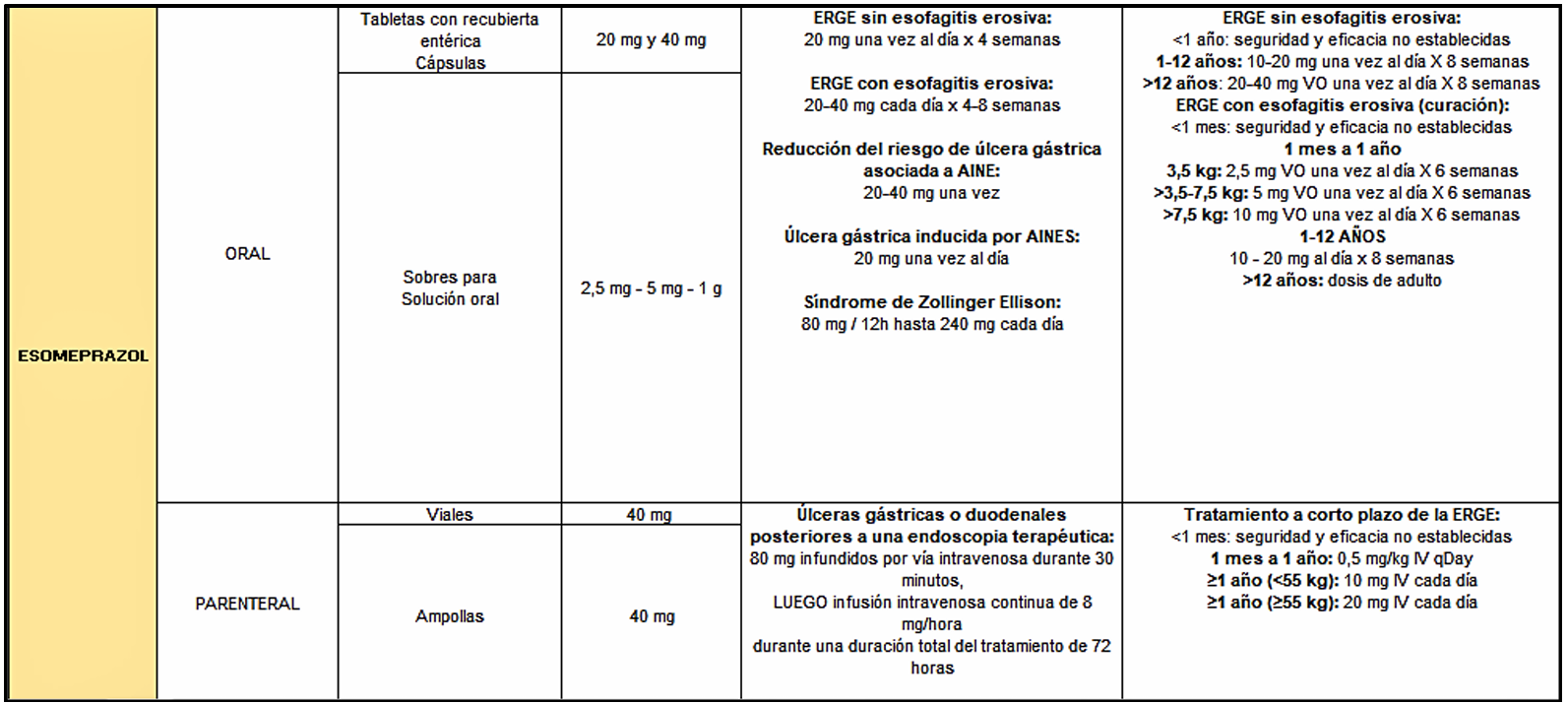
Los inhibidores de la bomba de protones (IBP), como omeprazol, pantoprazol, esomeprazol, lansoprazol, dexlansoprazol y rabeprazol, son profármacos que se activan en el medio ácido del canalículo secretor de la célula parietal [56].
Tras su absorción intestinal, se concentran en los canalículos donde el pH bajo induce su doble protonación, convirtiéndolos en formas activas (sulfenamidas o sulfenatos) que reaccionan con la H⁺/K⁺-ATPasa, enzima responsable del paso final de la secreción ácida gástrica [57].
Una vez activados, los IBP forman enlaces covalentes tipo disulfuro con residuos de cisteína del dominio luminal de la bomba de protones, principalmente en Cys813, aunque diferentes IBP se unen también a Cys822, Cys892 o Cys321, generando una inhibición funcionalmente irreversible hasta que se sintetizan nuevas bombas [58].
Aunque su vida media plasmática es corta, los efectos duran de 24 a 48 horas, ya que dependen de la velocidad de recambio enzimático y no de la concentración plasmática. Por ello, los IBP deben administrarse antes de las comidas, momento en que se activa el mayor número de bombas de protones [59].
Las diferencias estructurales entre IBP determinan variaciones en la estabilidad del enlace covalente y en la duración de la inhibición:
- Pantoprazol: se une a Cys813 y Cys822, formando complejos más resistentes a agentes reductores [60].
- Omeprazol y esomeprazol: se unen a Cys813 y Cys892 [61].
- Lansoprazol y dexlansoprazol: se enlazan a Cys813 y Cys321, con efecto similar [62].
Como consecuencia, la inhibición de la H⁺/K⁺-ATPasa disminuye la secreción de HCl, aumenta el pH intragástrico y favorece la cicatrización de lesiones ácido-relacionadas, como úlcera péptica o reflujo gastroesofágico [63].
¶ Conclusión
La producción de ácido clorhídrico en el estómago depende de la interacción coordinada entre mecanismos nerviosos, hormonales y paracrinos que garantizan una digestión eficiente y la protección contra microorganismos ingeridos.
Las vías estimuladoras, mediadas por la acetilcolina, la gastrina y la histamina, activan de forma conjunta la bomba H⁺/K⁺-ATPasa de las células parietales, mientras que distintos mediadores inhibitorios, como la somatostatina, las prostaglandinas y la secretina, limitan la secreción para evitar daños en la mucosa. Este equilibrio se ajusta de manera continua mediante la retroalimentación basada en los cambios de pH intragástrico.
Cuando este sistema regulador se altera, pueden desarrollarse enfermedades como la ERGE y las úlceras pépticas, lo que ha impulsado el uso clínico de los inhibidores de la bomba de protones (IBP). Estos fármacos reducen de manera sostenida la secreción ácida al bloquear la actividad de la H⁺/K⁺-ATPasa, facilitando la recuperación de la mucosa y el control de los síntomas. Comprender la fisiología de la secreción gástrica y la acción de los IBP es fundamental para optimizar el tratamiento de los trastornos relacionados con la hiperacidez y mejorar los resultados clínicos.
¶ Anexos
Fuente: Elaborado por los autores.
Fuente: Elaborado por los autores.
Fuente: Modificado por los autores. Adaptado de: Dreamstime.
Fuente: Modificado por los autores. Adaptado de: Dreamstime.
Fuente: Modificado por los autores. Adaptado de Instagram.
Fuente: Modificado por los autores. Adaptado de Google Share.
Fuente: Elaborado por los autores.
Fuente: Elaborado por los autores.
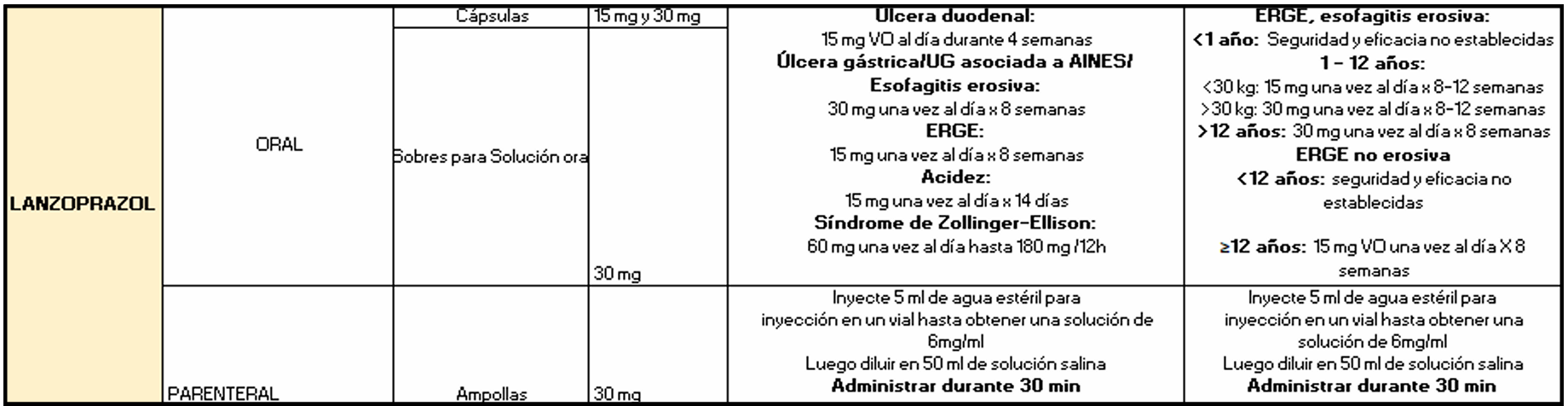
Fuente: Elaborado por los autores.
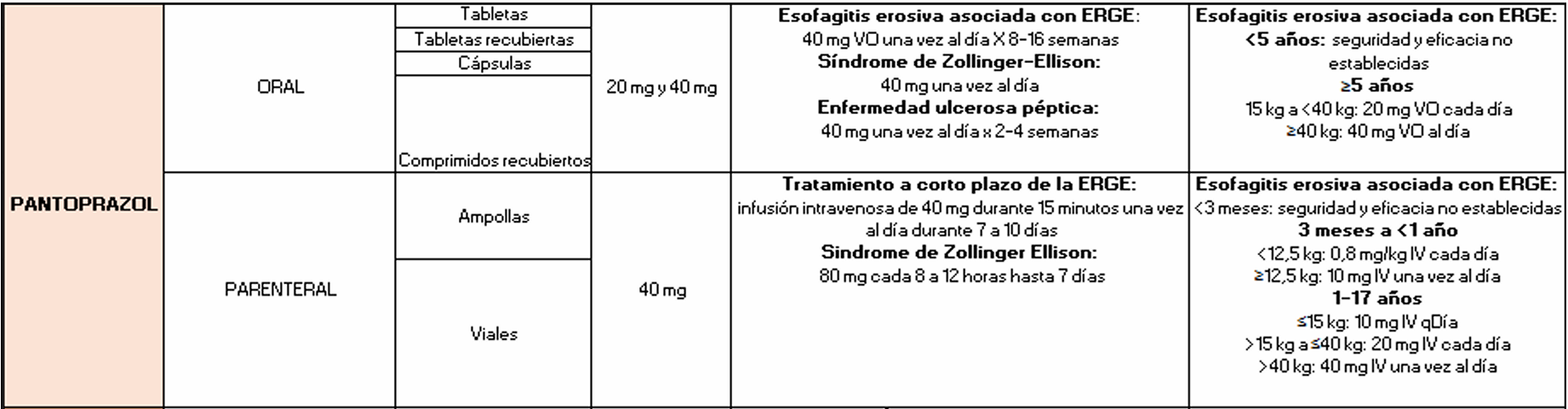
Fuente: Elaborado por los autores.

Fuente: Elaborado por los autores.
¶ Referencias
- Costanzo LS, Palacios Martínez JR. Fisiología. L'Hospitalet de Llobregat, Barcelona: Wolters Kluwer Health; 2015.
- Garza-Velasco R, Garza-Manero SP, Perea-Mejía LM. Microbiota intestinal: aliada fundamental del organismo humano. Educ Quím. 2021;32(1):10–19.
- Peña A. ¿Cómo funciona una célula?: fisiología celular. México: Fondo de Cultura Económica (FCE); 2016.
- Fabbiani S, Goyret A. Uso de inhibidores de la secreción ácida en adultos mayores. Bol Farmacol. 2023;14:1.
- Palacios Martínez JR, Preston RR, Wilson TE. Fisiología. L'Hospitalet de Llobregat, Barcelona: Wolters Kluwer Health; 2013.
- Cruz KAA. Química de alimentos. Ciudad de México: Universidad Nacional Autónoma de México; 2024.
- López Vallejo AB, González Jaramillo K, Valencia Paredes ML, Herrera Cedeño CM. Microbiota y su papel en el sistema inmunológico. RECIAMUC. 2022;6(2).
- Caballero-Barrios J, Caballero-Torres AE, Fernández-Vélez YE. Consideraciones breves sobre microbiota intestinal en la prevención y tratamiento de la COVID-19. QhaliKay Rev Cienc Salud. 2021;5(1):38–44.
- Castro E, Pérez F, Ramos A. Mecanismos de secreción de ácido clorhídrico: el rol de los transportadores iónicos en la membrana apical de la célula parietal gástrica. Rev Gastroenterol Mex. 2024;89(3):190–205.
- Shamsi BH, Chatoo M, Xu XK, Xu X, Chen XQ. Versatile functions of somatostatin and somatostatin receptors in the gastrointestinal system. Front Endocrinol (Lausanne). 2021;12:652363.
- Aguilar Salinas CA. Alexánderson: fisiología de los sistemas endocrino y digestivo. Ciudad de México: Editorial El Manual Moderno; 2019.
- Gonçalves TM. Ácido clorhídrico (HCl) en el estómago: propuesta de una clase práctica sencilla y económica para simular los aspectos fisiológicos de la digestión. Investig Soc Desarro. 2021;10(8):e20210816980.
- González Bustos L, Rodríguez Paz CA. Evolución del conocimiento y descripción del nervio de Latarjet en la anatomía y la cirugía. Cir Gen. 2025;47(2):113–118.
- Hsu M, Safadi AO, Lui F. Physiology, stomach. Treasure Island (FL): StatPearls Publishing; 2023.
- Varela CD, Segura N. Histology, parietal cells. Treasure Island (FL): StatPearls Publishing; 2023.
- Chen B, Liu G. Regulación integrada de la motilidad y la secreción gástrica: la visión del eje nervioso y endocrino en el estómago. Front Neurosci. 2023;17:1120000.
- Vernygorodskyi S, Martínez-Olivares M, et al. Gastric epithelial cell plasticity and molecular mechanisms of metaplasia. Gastric Cell Biol. 2025;6(3):24.
- Zhang X, Wang F, et al. Histamine secretion mechanisms from gastric ECL cells under physiological conditions. Front Physiol. 2021;12:708698.
- Pavlov VA, Chavan SS. Neural regulation of immunity and inflammation: implications for systemic diseases. Nat Rev Immunol. 2021;21(10):1–15.
- Sanger GJ. Muscarinic mechanisms in the upper GI tract. Neurogastroenterol Motil. 2022;34(2):e14243.
- Hattori T, Suzuki S, Tanaka M, et al. Histamine signaling in gastric physiology. Cells. 2023;12(5):812.
- Dockray GJ. Gastrin and gastric function revisited. Physiol Rev. 2022;102(1):169–210.
- Wallace JL. Prostaglandins and mucosal protection. Gastroenterology. 2023;165(1):30–45.
- Papantoniou K, Nikolaou M, Petropoulos K, et al. Somatostatin in gastrointestinal regulation. Biology (Basel). 2025;14(5):558.
- Laurila S, Laitinen K, Mäkelä J, et al. Secretin physiology revisited. Front Endocrinol (Lausanne). 2021;12:737686.
- Drucker DJ. Incretin biology from bench to therapy. Diabetologia. 2023;66:2541–2556.
- Chen D. Neuroendocrine regulation of gastric acid. J Neuroendocrinol. 2023;35(11):e13305.
- Hall JE. Guyton y Hall. Tratado de fisiología médica. Barcelona: Elsevier; 2021.
- Lee S, Park H. La interacción balanceada de los receptores de histamina y somatostatina en la célula parietal: el punto clave entre la digestión y la autoprotección. Gastroenterology. 2023;165(3):600-615.
- Buitrón R, Flores G. Regulación de la secreción ácida: un circuito trifásico integrado por vías neurales, endocrinas y parácrinas. Rev Fac Med (Méx). 2022;65(3):7-15.
- Salazar-López E, Torres-Ramírez F. Convergencia de señales estimuladoras en la bomba de protones gástrica. Neurociencia (Lima). 2024;7(2):110-125.
- Silva-García P, Ruiz-Díaz M. Identificación y función de los receptores estimuladores clave (M₃, H₂, CCK₂) en la célula parietal. Rev Chil Cardiol. 2023;42(4):300-315.
- Montes C, Ríos M. La activación vagal: un evento crítico en la fase cefálica de la secreción gástrica. Rev Cubana Med Intensiva Emerg. 2022;21(2):e1000.
- Romero S, Silva K. Localización y acoplamiento de receptores M₃ en la célula parietal. Rev Cubana Farm. 2023;57(2):e5208.
- Gómez J, Marín A. Vía de señalización Gq del receptor M₃: la cascada de IP₃ y DAG/PKC en la célula parietal. J Cell Sci. 2024;137(15):115-128.
- Martínez E. Los segundos mensajeros Ca²⁺ y DAG/PKC en la activación vagal de la célula parietal [Tesis Doctoral]. [Barcelona (ES)]: Universidad de Barcelona; 2023.
- Soto-Aguilar M, Vargas-López S. Mecanismos de inserción de la bomba de protones en la membrana apical mediada por Ca²⁺ y PKC. Endocrinol Nutr. 2021;68(5):340-355.
- Vega L, Castro R. Histamina gástrica: secreción por células ECL y su acción parácrina sobre la célula parietal. Rev Gastroenterol Peru. 2022;42(3):200-212.
- Soto A, Parra L. Receptor H₂: el acoplamiento a Gs y la generación de AMPc como señal estimuladora. Rev Gastroenterol Chile. 2022;47(1):5-15.
- López V, Guerra M. Cascada AMPc–PKA en la célula parietal: fosforilación y translocación de la bomba H⁺/K⁺–ATPasa. Rev Endocrinol Mex. 2025;14(1):120-135.
- Aguilera G. Gastrina y su función endocrina: unión a CCK₂ para la estimulación de la secreción ácida. Horm Mol Biol Clin Investig. 2023;5(1):12-25.
- Osorio-Alonso H, Ramos-Navarro C. Vía Gs del receptor CCK₂: similitud en la transducción de señales con ACh. Nefrol Mex. 2023;48(7):150-162.
- Butrón R, Flores G. Regulación de la respuesta inflamatoria: un nuevo rol del AMP cíclico en la célula parietal. Rev Fac Med (Méx). 2022;65(3):300-311.
- Pérez S, Rodríguez F. Mecanismos inhibitorios de la secreción de HCl: ejes esenciales para la homeostasis y la protección epitelial. Acta Endocrinol Mol. 2024;18(3):325-330.
- García J, Morales P. Acción pleiotrópica de los inhibidores: modulación directa e indirecta de las células clave (Parietal, G, ECL). Rev Chil Endocrinol Diabetes. 2025;18(1):50-65.
- Díaz R. Somatostatina gástrica: el inhibidor potente paracrino y endocrino de la secreción ácida [Tesis]. [Machala (EC)]: Universidad Técnica de Machala; 2024.
- Fernández L. La célula D como sensor de pH: liberación de SST y unión al receptor SSTR2 de la célula parietal [Tesis]. [Zaragoza (ES)]: Universidad de Zaragoza; 2024.
- Romero S, Silva K. Vía Gᵢ acoplada a SSTR2: la inhibición de la adenilato ciclasa como mecanismo de SST. Rev Cubana Farm. 2023;57(2):25–28.
- Pérez S, Rodríguez F. Doble acción inhibitoria de SST: control sobre la célula G y la célula ECL. Acta Endocrinol. 2024;18(3):25–30.
- Montes C, Ríos M. El papel de la Ciclooxigenasa en la síntesis de PGE₂ y su rol fisiológico gástrico. Rev Cubana Med Intensiva Emerg. 2022;21(2):e1000.
- Silva-García P, Ruiz-Díaz M. Receptor EP3 y su acoplamiento Gᵢ en la inhibición ácida mediada por PGE₂. Rev Chil Cardiol. 2023;42(4):300–315.
- Soto A, Parra L. Efecto citoprotector de las prostaglandinas. Rev Gastroenterol Lat Am. 2022;52(4):390–405.
- López V, Guerra M. Péptido inhibidor gástrico (GIP) y su origen intestinal (células K): doble acción sobre secreción y motilidad. REMUS. 2025;14(1):120–135.
- García J, Morales P. Inhibición de GIP: bloqueo de la liberación de gastrina y reducción de la actividad de la H⁺/K⁺-ATPasa. Rev Chil Endocrinol Diabetes. 2025;18(1):50–65.
- Martínez E. Inhibidores de la bomba de protones (IBP): efecto fisiológico y clínico. Gac Med Mex. 2023;159(4):450–456.
- Ahmed A, Clarke JO. Proton pump inhibitors. Treasure Island (FL): StatPearls Publishing; 2023.
- Wołowiec Ł, Szymański M, Wiciński M, Grześk G. Pharmacodynamics of PPIs. Front Pharmacol. 2025;10:1507812.
- Srebro J, Kurnik-Łucka M, Górska A, et al. PPI formulation and activation mechanisms. Pharmaceutics. 2022;14(10):2043.
- Andrawes M, Abdelmessih R, Hanna P, et al. Rational use and efficacy of PPIs. Medicina (Kaunas). 2025;61(9):1569.
- Shin JM, Kim N. Pharmacology of pantoprazole. Gut Liver. 2022;16(5):739–748.
- Andersson K. Omeprazole and esomeprazole binding profiles. Clin Pharmacokinet. 2021;60(7):877–890.
- Kukulka M. Dexlansoprazole pharmacology update. Expert Opin Drug Metab Toxicol. 2021;17(5):563–573.
- Moayyedi P, Eikelboom J, Bosch J, et al. Acid suppression and mucosal healing. Lancet Gastroenterol Hepatol. 2022;7(8):726–738.